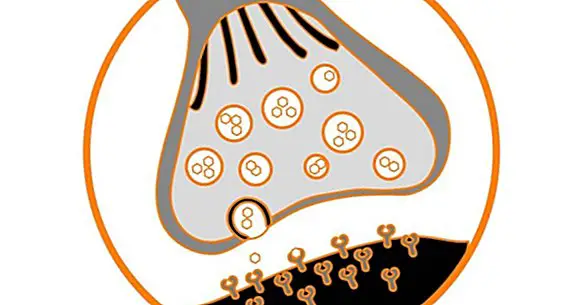
Synapsien historia
Aivot sisältävät tuhansia ja tuhansia yhteenliittymiä sen hermosolujen välillä, jotka on erotettu pienellä tilalla, joka tunnetaan nimellä synapseja. Siellä tiedonsiirto siirtyy hermosolusta neuroniin .
Jo jonkin aikaa on havaittu, että synapsin aktiivisuus ei ole staattinen, toisin sanoen se ei ole aina sama. Sitä voidaan parantaa tai vähentää ulkoisten ärsykkeiden, kuten asioiden, seurauksena. Tämän synapsin moduloinnin kyky tunnetaan nimellä aivojen plastisuus tai neuroplastisuus.
Tähän asti on oletettu, että tämä kyky moduloida synapseja osallistuu aktiivisesti kahteen toimintaan, jotka ovat tärkeitä aivojen kehitykselle oppimisen ja muistin kannalta. Sanon tähän asti, koska tässä selityksessä on uusi vaihtoehto, jonka mukaan ymmärtää muistin toimintaa, synapsien ei ole niin tärkeää kuten normaalisti uskotaan.
Synapsien historia
Ramón y Cajalin ansiosta me tiedämme, että neuronit eivät muodosta yhtenäistä kudosta, mutta kaikki ne erotetaan toisistaan interneuronaalisilla tiloilla, mikroskooppisilla paikoilla, jotka myöhemmin Sherrington kutsisivat "synapsiksi". Joitakin vuosikymmeniä myöhemmin psykologi Donald Hebb tarjosi teorian, jonka mukaan synapseja eivät aina ole samansuuntaisia ja niitä voidaan moduloida, eli hän puhui siitä, mitä tunnemme neuroplastiikaksi: Kaksi tai useampia neuroneja voi aiheuttaa niiden välisen suhteen vakauttamisen tai hajoamisen , jolloin tietyt viestintäkanavat ovat useammin kuin toiset. Kuten utelias tosiasia, 50 vuotta ennen tämän teorian soveltamista, Ramón y Cajal jätti todisteet tämän modulaation olemassaolosta hänen kirjoituksissaan.
Tänään me tiedämme kaksi mekanismia, joita käytetään aivojen plastisuuden prosesseissa: pitkän aikavälin potentiaatio (LTP), joka synapsin tehostaminen kahden hermosolun välillä; ja pitkän aikavälin masennus (LTD), joka on vastakkainen ensimmäiseen eli tiedonvälityksen väheneminen.
Muisti ja neurotiede, empiirinen näyttö kiistelyllä
Oppiminen on prosessi, jonka avulla yhdistämme asioita ja tapahtumia elämässä hankkimaan uutta tietoa. Muisti on toimintaa, jota ylläpidetään ja säilytetään ajan mittaan. Koko historian aikana on tehty satoja kokeita etsimään, miten aivot suorittavat nämä kaksi toimintaa.
Tämän tutkimuksen klassikko on Kandelin ja Siegelbaumin (2013) työ, jossa on pieni selkärangattu meri-etana, joka tunnetaan nimellä Aplysia. Tässä tutkimuksessa, he näkivät, että synaptisen johtokyvyn muutokset johtuivat eläimen reagoinnista ympäristöön , mikä osoittaa, että synapsi on mukana oppimisen ja muistiinpanon prosesseissa. Mutta viimeisimmän kokeilun Aplysia Chen et al. (2014) on löytänyt jotain, joka ristiriidassa aikaisemmin tehtyjen päätelmien kanssa. Tutkimus paljastaa, että elimistössä pysyy pitkäaikainen muisti moottoritoiminnoissa, kun synapsi on estynyt huumeiden avulla, epäilee ajatusta siitä, että synapsi osallistuu koko muistiprosessiin.
Toinen asia, joka tukee tätä ajatusta, johtuu Johansson et al. (2014). Tässä yhteydessä selvitettiin pikkuaisteen Purkinje-soluja. Näillä soluilla on tehtävänsä kontrolloida liikkeiden rytmiä, ja heitä stimuloidaan suoraan ja huumeiden synapsien eston alla ennustetta vastaan, he jatkavat vauhtia. Johansson totesi, että ulkoiset mekanismit eivät vaikuta muistiinsa ja että Purkinje-solut itse ohjaavat mekanismia erikseen riippumatta synapsien vaikutuksista.
Lopuksi Ryan et al. (2015) osoitti, että synapsin vahvuus ei ole kriittinen kohta muistin yhdistämisessä. Hänen työnsä mukaan tuotettaessa proteiini-inhibiittoreita eläimissä syntyy retrogradamnesia eli se ei voi säilyttää uutta tietoa. Mutta jos tässä samassa tilanteessa sovelletaan pieniä valonlähteitä, jotka stimuloivat tiettyjen proteiinien tuotantoa (menetelmää, joka tunnetaan nimellä optogenetics), voimme säilyttää muistin indusoitua kemiallista estämistä huolimatta.
Oppiminen ja muisti, yhtenäiset tai itsenäiset mekanismit?
Jotta voisimme muistaa jotain, meidän on ensin opittava siitä . En tiedä, johtuuko se tästä, mutta nykyinen neurotieteellinen kirjallisuus pyrkii yhdistämään nämä kaksi käsitettä ja kokeiluihin, joihin ne perustuvat, yleensä on moniselitteinen johtopäätös, joka ei salli erottaa toisistaan oppimisprosessia ja muistia, minkä vuoksi on vaikea ymmärtää, käyttävätkö he yhteinen mekanismi tai ei.
Hyvä esimerkki on Martinin ja Morrisin (2002) työ hippokampuksen tutkimuksessa oppimiskeskuksena. Tutkimuspohja keskittyi N-metyyli-D-aspartaatin (NMDA) reseptoreihin, joka tunnistaa neurotransmitterin glutamaatin ja osallistuu LTP-signaaliin. He osoittivat, että ilman pitkää potentiaalia hypotalamuksen soluissa on mahdotonta oppia uutta tietoa. Koe koostui NMDA-reseptorisalpaajien antamisesta rotilla, jotka jätetään vesirummulla lautalla, eivätkä pysty tutkimaan lautan sijaintia toistamalla testi, toisin kuin rotat ilman estäjiä.
Seuraavat tutkimukset paljastavat, että jos rotta saa koulutusta ennen inhibiittoreiden antamista, rotta "kompensoi" LTP: n menetykseen, eli sillä on muisti. Päätelmä, jonka haluamme näyttää, on se LTP osallistuu aktiivisesti oppimiseen, mutta se ei ole niin selvä, että se tekee tiedonhakuun .
Aivojen plastisuuden seuraus
On monia kokeita, jotka osoittavat, että neuroplastisuus osallistuu aktiivisesti uuden tiedon hankintaan , esimerkiksi edellä mainittu tapaus tai siirtogeenisten hiirten luominen, jossa glutamaatin tuotannosta poistetaan geenit, mikä vaikeuttaa eläimen oppimista.
Sen sijaan roolisi muistissa alkaa olla epäilemättä, kuten olet lukenut muutamalla esimerkillä. Teoria on alkanut nousta esille, että muistin mekanismi on solujen sisällä pikemminkin kuin synapseissa. Mutta kuten psykologi ja neurotieteilijä Ralph Adolph osoittaa, neurotiede ratkaisee, miten oppiminen ja muisti toimivat seuraavien viidenkymmenen vuoden aikana , eli vain kerran selvennetään kaikkea.
Kirjallisuusviitteet:
- Chen, S., Cai, D., Pearce, K., Sun, P.Y.-W., Roberts, A.C. ja Glanzman, D.L. (2014). Palautetaan pitkäaikainen muisti sen jälkeen, kun sen käyttäytymis- ja synaptiivinen ilmaisu poistettiin Aplysiaan. eLife 3: e03896. doi: 10.7554 / eLife.03896.
- Johansson, F., Jirenhed, D.-A., Rasmussen, A., Zucca, R., ja Hesslow, G. (2014). Muistin jäljitys ja ajoitusmekanismi, joka on lokalisoitu pienisillä Purkinje-soluilla. Proc. Natl. Acad. Sci. U.S.A. 111, 14930-14934. doi: 10,1073 / pnas.1415371111.
- Kandel, E. R. ja Siegelbaum, S. A. (2013). "Immlusiivisen muistin varastointimekanismit ja yksilöllisyyden biologinen perusta", Neural Science -periaatteet, 5. Edn., Eds ER Kandel, JH Schwartz, TM Jessell, Siegelbaum SA ja AJ Hudspeth (New York, NY, McGraw-Hill ), 1461-1486.
- Martin, S.J. ja Morris, R.G. M. (2002). Uusi elämä vanha idea: synaptinen plastisuus ja muisti hypoteesi tarkistetaan. Hippocampus 12, 609-636. doi: 10.1002 / hipo.10107.
- Ryan, T.J., Roy, D.S., Pignatelli, M., Arons, A., ja Tonegawa, S. (2015). Engram-solut säilyttävät muistia retrogradamnesiassa. Tiede 348, 1007-1013. doi: 10.1126 / science.aaa5542.